Halo of Confusion: Why Some Smart People Entertain Darwinian Doubts (Part 2)

Photo by Michael Dam on Unsplash
27 minutes
In the conclusion to Part 1, I said that I would move from abstract logical critiques of Neo-Darwinism, to a critique of the concrete evidence it generally offers for the efficacy of evolution by mutation + natural selection. From there I would move to an analysis of why Darwinists generally showcase the worst, rather than the best evidence for that fact that some kind of evolution has taken place. But before I get there, I want to address one more abstract issue: Darwinian reductionism.
I. A Universal Acid
Consider the sense of wonder evoked by one of our modern nature documentaries. Any good nature show is like a story. It carefully establishes the setting, before narrowing its focus to one or more of the players, who are then given a good description; their unique characteristics, their abilities, their needs, their virtual personalities, etc. Then comes the inciting incident that sets the stage for the drama. More often than not, this a predation scenario, though modern biology is certainly more aware than were naturalists in Darwin’s time of the widespread forms of mutualism, cooperation, and near-altruism in nature. But despite this already one-sided emphasis on competition, the show still ensnares its viewer in a gripping story. In doing so, it draws him upwards and outwards, toward the dance of creation, and toward the mystery of being; of goodness, truth, and beauty considered for their own sakes. Like a game, a poem, or a prayer, this approach to nature is fundamentally whole-some, because it puts the soul in contact with unique substantial forms, and their specific, fascinating interrelations. With the whole dance of life. But then comes Darwin…
Sooner or later, the beauty and mystery above will be boiled down to an ugly, primordial scuzz. We the viewers will learn, via Darwinian analysis, that this organism’s specific, defining characteristics are but adaptations – though of what previous feature we’re never told. That glorious colored parrot chewing clay to get its nutrients ceases being a thing with specific qualities, and becomes adaptation piled upon adaptation, joined together with other adaptations. Soon there is no longer a parrot at all, only a network of connected relations; slow motion changes of an almost-something into an almost-something-else. The parrot itself disappears in the analysis of parrot, and we are still only on the level of morphology. And morphology itself is, on the Neo-Darwinian analysis, only a kind of illusion, an external raiment or carrier for the real drama, which is the drama of genes. And genes are but chemicals in certain stable arrangements. They are “selfish” replicators, wearing beautiful, phenomenological disguises, which we, who are subject to our own adaptation-induced illusions, mistake for meaning. Meaning itself is but an adaptation, a survival strategy of mindless replicators who have produced consciousness as an epiphenomenon of their inexorable, grinding, chemical competition.
Now while conceding that an organism’s unique characteristics must, generally speaking, be adaptive, there is a logical problem with referring to them in the nominative case as “adaptations.” The features of a particular bird are also definitional of what that species of bird is. It makes no sense to say that the wings of a parrot are adaptations that allow it to fly. The “it” there refers to “parrot”, and wings (or the remnants of wings, in some cases) are definitional properties of birds. One might as well say that a car’s wheels are “improvements” that allow it to drive. A car, before the “improvement” of wheels, simply isn’t a car! In the physical world, there is no such thing as a “general bird,” simple and un-adapted to its environment. Everything that lives necessarily has features that permit it to live. These features are adaptive, but to always call them adaptations is just to read transience into essence. I realize that this objection to the constant use of the word adaptation to mean “feature” might seem pedantic, but there is something truly maddening and reductionist about this use of language.
When philosopher Daniel Dennett approvingly called Darwin’s idea a “universal acid” eating through all of our conceptions of meaning, morality, and even truth, he was only being reasonable. The Darwinian mode of analysis, though it begins with whole organisms, tends towards a severe reductionism. The appeal of this sort of analysis for the secularist is that, like every form of rebellion, it purports to tear down one form of dogma, but ends by transferring power into the hands of the revolutionary. And if we mere laypersons are so deluded by our un-scientific sense of goodness, beauty, truth — and thing-ness! — and yet, if we cannot possibly live without those same transcendentals, then we shall need an interpreter, one of those wise beings who, like Dr. Dennett, has exchanged the garb of a white wizard for a cloak of many colors. There will, we are told, be a few compromises – especially when it comes to personal morality, and our sense of human dignity – but never fear; these new high priests will give us new causes and new rituals which, when embraced with existential courage, may be almost as meaningful to us as the old meanings were in themselves. Have we lost the sense that there are things, and that we can see, know, and name them? Never fear! Saruman Daniel Dennett is here!
An Antidote
How do we refute this corrosive pattern of thought, particularly if – despite doubts concerning Darwin’s mechanism – we find common descent likely anyhow? How, in other words, are we to think about a pattern of change over time, without change itself becoming the only reality? I would like to suggest a few ground rules which can help us to navigate these confusing metaphysical waters without being consumed by them.
First, consider that everything we can infer or deduce about the history of life starts with real organisms. Let us take a common bird for an example: the American robin. Now the robin is a kind of thrush, or is closely related to thrushes, but whatever else it may be, it is, first and foremost, itself. It has a certain characteristic body, coloration, diet, call, and set of behaviors. Its bill shape, feathers, method of flight, and seasonal patterns are stable and recognizable. When two robins mate, they produce offspring that are also robins, and whose qualities fall within a certain definable range. Within the species of American robin there are seven subspecies, the borderlines between them being somewhat vague, as might be expected of varieties within the same race.
As for the robin’s ancestry, Wikipedia says the following:
A study of the mitochondrial cytochrome b gene indicates that the American robin is not part of the Central/South American clade of Turdus thrushes; instead it shows genetic similarities to the Kurrichane thrush (T. libonyanus) and the olive thrush (T. olivaceus), both African species.[8][9] This conflicts with a 2007 DNA study of 60 of 65 Turdus species, which places the American robin's closest relative as the rufous-collared thrush (T. rufitorques) of Central America.
Well, we’ll take it as a given that the American robin is related to something, and that it came from somewhere. The difficulty for us is how to go about thinking about this pattern of change, whatever it may have been. At some point, surely, the American robin had an ancestor that was not a robin. For that matter, the first definable thrushes were preceded by some earlier birds that did not meet all the criteria of thrush–ness. But how are we to think about such a pattern of change without losing all sense of definite substances possessing definite accidents? How are we to escape Darwin’s acid bath?
The answer I propose it to climb into our mental time machine, and hop back through the robin’s ancestry. At some point, we’ll find a bird that has some but not all of the American robin’s defining characteristics. There are several possibilities here, depending on how far we go back, and we need to understand how to think about all of them. Suppose that at ten thousand years we find a creature with the same behaviors, seasonal patterns, plumage, and basic body design, yet at only half the size our own American robin. In that case, we still have the same species in an ontological, but not a biological sense, since the difference is only one of size. Going back ten thousand years more we find another ancestor, this time with different coloration and now a more restricted diet. Perhaps its seasonal patterns are also significantly different. Is it still a robin? Is anything anything? Do words have referents? Does evolution not reveal the world to be a kind of ontological kaleidoscope without reference points?
This is where we must calm down, and use our reason. We must remember that words like “robin” are our labels for things, and that the things themselves are the real substances. The word “robin” refers to a particular, definable organism that exists within a certain range of time. If ancient humans knew some other bird, ancestral to our own robin, they would also have a name for it, and would know its specific, defining characteristics. That substance, that bird that they knew, was a certain definable thing, and our own robin is also certain definable thing. Each stable species in the intervening time was a certain definable thing, and each stable species preceding it was a certain definable thing. If we calm down, and think clearly, we shall see that there is no morphological kaleidoscope. The kaleidoscope is an illusion produced by our imagination. The birds themselves are the real things.
The problem is that it is difficult for human beings to think about becoming. As a grammar teacher, I have reason every year to explain that “becoming” is a being verb, not an action verb. Grammatically, the sentence “The boy is John” is very much like the sentence, “The boy became a man.” Both “John” and “man” are predicate nominatives, nouns that follow a being verb and name or define the subject. This is because “boy” and “John” are the same entity, and “boy” and “man” are the same entity. The linking verbs “is” or “became” function to predicate a relationship of identity between the subject and the subjective complement.
The Law of the Excluded Middle states that between being and non-being there is no middle state. An entity either exists, or it doesn’t exist. Becoming is not an entity, but a succession discrete states, each of which states refers to some specific entity. In the case of evolution, then, we must remember that there is no slushy “becomingness” which constitutes the corrosive hidden “truth” about organisms. Instead, there is a succession of definite things.
Of course our words about those things will be somewhat artificial and imperfect, but that was already the case before evolution entered Western thought. One does not have to be a nominalist or a conceptualist to concede that, as Aquinas said somewhere, the human mind cannot fully comprehend the essence of even a single fly. If we cannot fully comprehend it, then neither can we perfectly name it. There is a certain imprecision in our names and thoughts about the substances we encounter. But things are still things. If they were not, we would never be able to talk about them. We certainly would not be able to infer evolution from the pattern of their succession, unless that succession were composed of actual entities. So much then for the universal acid. There can be no “flux” without things to fluctuate, and those things, not the imaginative impression of flux, are the real object of human thought.
II. Non-Evidence and Bad-Evidence
With these considerations in mind, we turn our focus to the poor evidence offered as proof of common descent. Strangely, it is also the most common evidence offered. It’s a bit of a mystery that Darwinists put their worst case first, and practically bury their best cases, but we have a theory about that as well, and will save it for the end. In the meantime, here are the most common textbook proofs, with an explanation of why most of them prove little or nothing.
Homology as Proof of Common Descent
We’ve already discussed the circularity using homology as evidence for common descent, while simultaneously proposing common descent as the explanation for homology. We also pointed out that there’d be no problem with proposing that common descent causes homology, provided that we defined the latter independently of the former. It turns out, however, that the issues of circularity noted above are not purely academic. They go right to the core of Darwinian reasoning, and constitute its biggest evidential blind spot.
Suppose we accept that homologies – shared biological features – are strong evidence for shared ancestry. If this is true, then we ought to find that highly similar structures only occur in believable lines of descent. Yet that is far from the case. There are numerous instances in the history of life of structures that appear to be homologous but which show up in lines that cannot possibly be closely related. This happens at both the morphological and molecular levels. Look, for example, at the skulls of the recently extinct thylacine (Tasmanian Wolf), and the grey wolf. To a layperson, the two are hard to tell apart. Certainly if the thylacine’s skull had been found as a fossil in the same area in which grey wolves currently live, we would expect the former to be identified as a progenitor of the latter. But the Tasmanian wolf was a marsupial, a member of an entirely different and geographically isolated group of mammals, and living at the same time (on a totally different part of the earth) as the grey wolf.

Since, in this case, we have strong independent evidence that the thylacine is not closely related to the grey wolf, the homologous appearance of these skulls cannot be interpreted as evidence of shared ancestry. In cases like these – and there are many – evolutionary biologists point out, reasonably enough, that evolution has independently arrived at a very similar structure. Similar “selection pressures” produced similar outcomes, though with only random mutation to provide the raw material. Leaving aside, for the moment, the intriguing fact that evolution “found” the same structure twice, or the improbability that the correct “random” mutations would be able to provide the raw material needed, let us consider how damning this is for the argument that homologous structures constitute evidence for shared ancestry.
Clearly, homologies prove no such thing, for their virtual equivalent can be produced by convergent evolution. And this destroys the whole principle upon which homology-as-shared-ancestry rests. The best one can say then is that shared homology is consistent with shared ancestry, except when it isn’t. But if it isn’t in this case, then how do we know it ever is? How, without the kind of pristinely preserved sequence of change that is rare in the fossil record, can we really be certain that homologous structures prove an ancestral relationship, as opposed to convergent evolution?
The matter of homology and analogy becomes even more distressing at the molecular level. Consider the eyes of a mouse, and the compound eyes of a fruit fly. Now these structures are not even homologous, but only analogous; that is, they are two fundamentally different structures that perform a similar function. But as this article relates, the same Pax-6 gene can be swapped between these vastly different organisms, even after their own respective eye genes are removed, and can be made to induce the species-correct eye for each.

To try to understand to bizarrity of this situation, imagine removing the genes for a turtle’s shell, and producing a line of shell-less turtles. Now take the embryo of one of these shell-less turtles, and insert the genes for a snail’s shell. Now suppose the turtle develops normally, making use of the snail-shell gene to produce an entirely normal turtle shell! That is the level of weirdness we are talking about! It is certainly suggestive of something, even perhaps of a certain evolutionary logic, but (contrary to Walter Gehring’s labored rationalization in the linked article) it does not not sound very Darwinian.
The Universality of DNA
Another problematic argument offered for common descent is the universality of the genetic code. That all organisms on earth utilize the same molecular toolkit is supposed to show that they must all be related by ancestry. But a few moments consideration reveals that the universality of DNA does not prove common ancestry. To see what I mean, imagine that scientists were to find, in some deep volcanic vent, a strange organism without DNA. We will suppose that this creature has some other, simpler genetic coding system, built on only three nucleotides instead of four. Now do you suppose for one moment that evolutionists would regard this hypothetical organism as evidence against common descent? If you have any familiarity with this breed of scientist, then you know that they would concede nothing of the kind. Instead there’d be a just-so story to the effect that this odd creature represents the descendant of an isolated, primitive sort of organism whose genetic code was a primitive precursor to DNA. We’d hear clever tales about how its descendants most likely gained a fourth nucleotide, resulting in vastly greater opportunities for generating biological diversity, etc. Indeed, its three nucleotide system would be hailed as just the sort of stepwise, jury-rigged process that evolution predicts.
Now the point here is not that the universality of DNA is inconsistent with common descent. The point is that it is merely consistent with it. Common descent would not be disproven by the discovery of some ancient remnant of a pre-DNA past. Nor is it proven by the actual universality of the genetic code. Either scenario could be consistent with – or inconsistent with – common descent. A piece of evidence that can go either way does not prove anything.
Moths and Finches
One of the prototypical evidences of natural selection in action is industrial melanism in peppered moths. During the Industrial Revolution, when air pollution became a serious problem, the more common white and black variety of peppered moth was superseded by the darker variety. This is supposed to be a dramatic example of nature “selecting” one form – the more naturally camouflaged form – over another.
Now this may be a fine example of the principle of natural selection, but it falls very short of showing that natural selection has produced all varieties of life on earth. Both forms of Biston betularia existed prior to the Industrial Revolution. During this period when black soot covered many surfaces upon which the moths would tend to perch, the less common black form became the predominant variety of peppered moth, as its coloring provided better camouflage against predators. When circumstances changed, the speckled white and black form again became predominant, as its coloring favored it in the new, cleaner environment.

But this is merely a shift in the frequencies of already existing animals. It represents a very weak form of microevolution, but does not even show how the different forms of peppered moth appeared in the first place, let alone how moths, butterflies, etcetera, appeared on earth. Indeed, even if a future study were to find that the shift from white to black was actually an epigenetic change – a morphological change activated from the outside in by as yet little understood mechanisms – such a change would also assume the preexistence in the moth’s genome of both coloration options. Likewise, if a study were to show this occurred by random mutation, which would, at best, show that a superficial characteristics can be altered by mutation. In other words, none of these possibilities get at the question of how genuine novelty appears in the history of life. Darwinists know this, of course, but seem happy to employ industrial melanism as a dramatic instance of evolution in action — as long as no one objects.
Haeckel’s Fraud
Another standard textbook proof for evolution are various drawings (or photographs) of vertebrate embryos which purport to show a similar path of development for related organisms. Ernst Haeckel introduced the notion that ontogeny (pre-birth development) recapitulates phylogeny (passes through the stages of evolutionary development.) Haeckel made this point by producing a series of famous woodcuts that seem to show that mammals, reptiles, fish, and the like go through an analogous process of fetal development, which process images their evolutionary history.

Now the problem here is two fold. First, Haeckel’s drawings were well-known frauds. Second, even photographs purporting to show the same relationship involve careful selections of just those embryos that seem superficially to demonstrate the idea that embryonic development shows evolutionary history. As this article notes, even major evolutionary figures have criticized the prevalence of Haeckel-like drawings in textbooks. But if Darwinists know that “ontogeny recapitulates phylogeny” is a false concept, why does this idea (or a near equivalent) seem to pop up over and over again in public demonstrations of evolution? Perhaps because, like industrial melanism, they can get away with it? Or is it because the version of evolution that this suggests is more consonant with Darwinism?
Germ Evolution
The capacity of diseases and viruses to rapidly mutate gives us a more dramatic and useful example of microevolution than do fluctuations in existing moth and finch populations. In the cases of germs, real time changes in the genetic makeup of the germ result in morphological changes, and gained or lost abilities. Because microorganisms multiply so rapidly, variations appear much more quickly. Those new variants for which the immune system or competitor isn’t prepared then multiply rapidly, outcompeting the parent stock.
While germ evolution does provide ready proof of micro-evolution – horizontal changes within an existing family of organisms – it does little to establish the more grandiose Darwinian claims. For one thing, the diseases and viruses in question obviously remain diseases and viruses, and of the same basic kind, after a variant population has emerged. For another thing, a mutation that leaves a germ resistant to antibiotics (for example) often involves a loss of function, a devolution in the program that results in structural changes that make the pathogen weaker. What is missing in the Neo-Darwinian story is any mechanism for producing fundamentally new information, new body plans, or a new direction for evolution. And that is a huge problem, for, as we’ve previously noted, mutation has to bear the entire burden of novelty.
As for microorganisms becoming something other than microorganisms, there do appear to be processes of symbiosis in the history of life whereby “simple” organisms have merged to form higher level organisms. However, it is interesting to note that the late Lynn Margulis, the lead proponent and originator of this symbiotic approach, was also a serious critic of the Neo-Darwinian explanation of evolution, as seen in the quote below:
“We agree that very few potential offspring ever survive to reproduce and that populations do change through time, and that therefore natural selection is of critical importance to the evolutionary process. But this Darwinian claim to explain all of evolution is a popular half-truth whose lack of explicative power is compensated for only by the religious ferocity of its rhetoric. Although random mutations influenced the course of evolution, their influence was mainly by loss, alteration, and refinement. One mutation confers resistance to malaria but also makes happy blood cells into the deficient oxygen carriers of sickle cell anemics. Another converts a gorgeous newborn into a cystic fibrosis patient or a victim of early onset diabetes. One mutation causes a flighty red-eyed fruit fly to fail to take wing. Never, however, did that one mutation make a wing, a fruit, a woody stem, or a claw appear. Mutations, in summary, tend to induce sickness, death, or deficiencies. No evidence in the vast literature of heredity changes shows unambiguous evidence that random mutation itself, even with geographical isolation of populations, leads to speciation. Then how do new species come into being? How do cauliflowers descend from tiny, wild Mediterranean cabbagelike plants, or pigs from wild boars?” LYNN MARGULIS, DORION SAGAN, ACQUIRING GENOMES: A THEORY OF THE ORIGINS OF THE SPECIES, (BASIC BOOKS, 2003), P. 29.
Incidentally, quotations like those above could be multiplied almost ad infinitum, as evolutionary biologists are often forced to confront the very un-Darwinian realties of the history of life, and of molecular biology. Neo-Darwinists close ranks, and put on a strong face, when speaking to the public about the truth of evolution by random mutation and natural selection, but many of them privately doubt that this is how evolution works. They know well that the textbook proofs – the same evidences endlessly repeated in science programs – are no good. These kinds of proofs do not touch the main point, which is how anything genuinely new appears in the history of life.
But if this is the case, then what are the good reasons – if any – for believing in the general fact of macroevolution/common descent, and why are Neo-Darwinians so quick to group criticism of their doctrine with the affirmation of young earth creationism, or pseudo-science? As I’ll argue below, these two questions have related answers.
Creationism as Neo-Darwinism’s Necessary Foil
Among the many people, secular and religious, who’ve expressed serious doubts about Darwinism, a minority are fundamentalist Protestants who believe in a young earth. These are the folks who argue that the earth is only a few thousand years old, and that its entire history lines up, more or less, with the Old Testament history from Adam on. Given the overwhelming evidence of entire worlds of extinct life forms, young earth creationists (YECs, hereafter) are forced to hold that Adam and Eve were contemporaries with the dinosaurs, and that the extinction of so many ancient organisms were caused by the Deluge. Embracing this position involves disregarding or reinterpreting vast swaths of evidence in the geologic column for a succession of life forms laid down over vast periods of time. It also involves ignoring or reinterpreting the evidence of astronomy, physics, and chemistry, all of which converge on a multi-billion year-old earth in a multi-billion year-old universe.
I will not do much to refute the YEC position other than to say that it is as unsustainable as the idea of a flat earth. There is simply too much overwhelming evidence that the earth and universe are much, much older than the biblical timeline, for this position to possibly be true. To invest the time to refute it would be to go down endless rabbit holes for the very small reward of discovering that a highly unlikely thesis was in fact false. Indeed, the situation here parallels the situation of a Catholic confronted by an evangelical who claims (as once happened to this author) that, “the early Christians were all Evangelicals and Pentecostals!” That claim is so overwhelmingly and completely false, so much in direct contradiction of literally all of the evidence of the writings of the Church Fathers, that the thought of trying to refute it is positively anxiety inducing. Literally everywhere you look in the early Church, you find references to the Eucharist, apostolic succession, the Catholic Church, the primacy of the bishop of Rome, Mary as the Second Eve, etc. In the same wise, literally everywhere you look in the fossil record you find overwhelming evidence of an old earth, and of a long succession of different organisms.
But because young earth creationism is so wrong, it makes a perfect foil for Neo-Darwinism. As mentioned in the Lynn Margulis quote above, Neo-Darwinists simply cannot make the case that random mutations provide the vast creative potential upon which natural selection can “go to work.” Knowing their position to be weak, they strike out with religious fervor against something they call “creationism.” To listen to their rhetoric, you would think that the only people who disbelieve in the Neo-Darwinian mechanism are fundamentalists. In Darwinian writings, all critics of this implausible explanation of life are grouped into the same camp.
And there’s good strategic reason for this. When one’s own position is weak, and one is not willing to concede its weakness, it makes sense to deflect. And the same logic of deflection holds true for the YECs. Thus the Neo-Darwinists and YECs are natural foils of each other. They are both in perfect agreement that there are only two options: undirected evolutionary processes, or a vulgar and obvious form of creation. With this binary established, Darwinists need only show the absurdity of the latter in order to legitimize the former.
But this strategy also begins to suggest an answer to the question I posed at the beginning of last week’s blog post: Why do Darwinists lead with the worst evidence for evolution? To discover the answer, we must first look at a few of the best arguments for some kind of macroevolutionary history of life, and then see how what they suggest differs from what Darwinism would predict.
III. The General Case for Evolution
The Pattern of Life History
It might have been the case that all of the different kingdoms of life, with their various orders, genera, and species, appeared at the same time in the past. Were that so, the only reasonable explanation would be a sudden, singular act of special creation, one in which secondary causality played a minimal role. But this is not what the fossil record shows.
Instead, the simplest cellular life appears first (almost as soon as the earth cooled,) followed by long periods in which there is only cellular life, followed by the geologically abrupt appearance of more complex microbial life, followed by the appearance of weird, more complex organisms, followed by the Cambrian Explosion, in which members of nearly all of the existing phylum on earth appear within the same geological window. From there, over vast stretches of time, new organisms appear within those existing phyla. Among vertebrates (in phylum chordata) fish appear before amphibians, which appear before reptiles. Certain reptilians appear in the fossil record which show skeletal features that are usually only seen in the later-appearing mammals. Other branches of reptilians show bird-like features. True mammals and true birds follow their reptilian intermediates, and then continue to live alongside reptiles. Within the order of mammals, one finds ancient mammals (such as sloths, horses, and elephants) which are like and unlike their modern counterparts. Some kind of progression of forms is on display here. Finally, among the order of primates, a genus of upright walking primates emerges. Among the species in this hominid genus, creatures physically similar to man show up in the fossil record. Finally a sub-species of homo sapiens emerges. Among this sub-species, at some point tens of thousands of years ago (perhaps 45,000 years ago; perhaps earlier) there is a recognizable leap forward to a creature who is clearly one of us.
Now the latter pattern unambiguously shows a progression of life. In general, simple things precede more complex things, and current organisms are anticipated by similar but extinct organisms, which once occupied the same geographical regions. This pattern appears on its face to be one of ancestral descent, because the latter organisms seem to form a trajectory of form and family from the earlier organisms. There are even creatures, like the mammal-like reptiles and the bird-like reptiles and an ancient amphibian with both lungs and gills, which are intermediate in structure between distinct groups, and whose fossils appear in strata that are intermediate in time between the two distinct groups. All of this strongly suggests a pattern of descent, and of new organisms somehow emerging from certain lines of existing organisms, and only in specific regions. True, the pattern is broken at many points, but its general shape is clear.
Evidence from Retroviruses
Certain viruses leave a fingerprint in the DNA of those organisms infected by them, and that fingerprint is passed on to their offspring. Talkorigins notes:
“Retroviruses (like the AIDS virus or HTLV1, which causes a form of leukemia) make a DNA copy of their own viral genome and insert it into their host's genome. If this happens to a germ line cell (i.e. the sperm or egg cells) the retroviral DNA will be inherited by descendants of the host. Again, this process is rare and fairly random, so finding retrogenes in identical chromosomal positions of two different species indicates common ancestry.”
Now it is a fact that human beings and chimpanzees have the same retroviral fingerprints in the exact same place in their genomes. There is simply no good explanation for how it could be there unless humans and chimpanzees shared a common ancestor. I also ask the reader to note how much stronger this is as a piece of evidence than the universality of DNA. All organisms having DNA is logically consistent with a number of different scenarios, including the special creation of each different originating line of organisms, with the creator simple employing the same system. However, man and chimpanzees having the same retroviral imprint in the exact same place in their respective genomes only makes sense if they share the same ancestor, which at some point contracted that virus. We may imagine a teacher leaving a false answer key on his desk because he suspects cheating, and then finding that several students have copied those exact incorrect answers in the same exact order on their tests.
Evidence from Biogeography
Talkorigins notes:
“A reasonable nonevolutionary prediction is that species should occur wherever their habitat is. However, macroevolution predicts just the opposite — there should be many locations where a given species would thrive yet is not found there, due to geographical barriers.”
This is well-stated. Unless organisms developed and emerged in particular times and places, there would seem to be no good reason for these organisms, or others that replace them when they go extinct, to be naturally found only in those regions. After all, many organisms can thrive far from where they naturally occur on the globe. There are no native placental mammals in Australia, for example, but rabbits and dogs thrive there. The same is true of many plant species, which appeared in only one or a few proximate regions, but which can thrive in foreign environments. On the other hand, seemingly out-of-place creatures (like American marsupials) can almost always be shown to have originated elsewhere, when continents were still joined together.
While biogeography does pose some difficulties for common descent – the geological timing for monkeys in South America does not seem to work — on the whole it still supports the idea of common descent. Biogeographical distribution looks arbitrary apart from common descent, but makes perfect sense in light of it.
Convergent Evolution
The last general proof we’ll look at for evolution is the widespread phenomenon of morphological convergence. By this we mean the exceedingly strange fact that organisms that cannot possibly be related often have similar structures. There are many examples of this, but I would like to focus on the most breathtaking examples: the convergence of marsupial and placental mammal forms.
Think about this for a moment: There are placental mice, and marsupial “mice.” There are placental flying squirrels and marsupial flying “squirrels.” We’ve already seen the shocking similarity of skulls of placental and marsupial wolves. Would you believe me if I told you that in addition to the extinct saber-toothed tiger (placental) there was also a marsupial saber-toothed “cat”? Well there was!
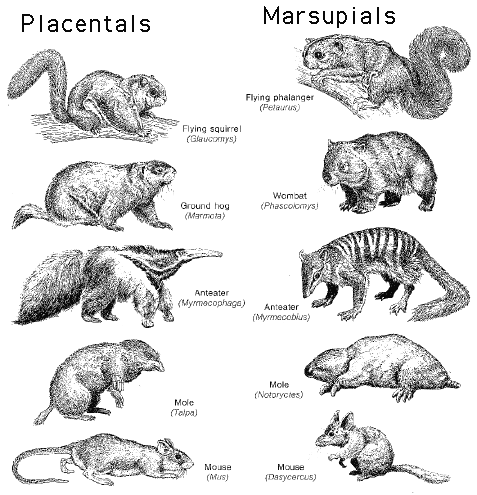
Now, on the one hand, when combined with biogeography, and with the fossil evidence that shows a prehistory of marsupial (and monotreme) development, convergence makes a powerful case for evolution, in light of geographical isolation. Why should these “alternative” forms appear over time on an isolated continent, unless they developed there? But on the other hand, it is simply impossible for any reasonable person to look at this pattern of morphology – and the way it parallels the trajectory of placental morphology – without seeing clear evidence of a plan. Of some kind of deep design logic rooted in the nature of life, and perhaps of matter itself.
How, you might wonder, do Neo-Darwinists, committed as they are to the idea that evolution is aimless, explain this weird parallelism? They do so by claiming that similar forms were produced by similar “selection pressures.” In other words, while the mutations are still random, the environmental circumstances that “shape” organisms are not random. Natural selection, the fact that animals survive if they have survivable features, is supposed to be a delicate enough instrument to create flying squirrels twice! I say not on your life.
Remember that under Darwinism mutation must bear the entire burden of creating novelty. Remember also that to use the word “mutation” to mean simply “whatever it was that happened such that we get the new genomic stuff we need for the theory to be true” is to simply beg the question. Finally, remember that mutations – copying errors – are supposed to be random. Therefore, before any “selection pressures” can “go to work” there must first emerge totally by accident all of the helpful genomic novelties which are subsequently blindly shaped into things like wolves, mice, flying squirrels, anteaters, and groundhogs, and saber-toothed cats. And these same one-in-a-million copying errors must happen over and over again for millions of years on entirely separate continents with exceedingly similar outcomes. No wonder Darwin and his followers rhapsodize about natural selection as if it were a kind of meta-mind! If there were no God, Darwinists would have to invent Him.
IV. The Real Reason Darwinists Lead With Weak Arguments
At the beginning of the first article, I promised to offer my thoughts on why Neo-Darwinists prefer to offer their worst evidence, rather than emphasizing the best evidence for evolution, with the result that many careful thinkers just don’t find them credible, with the further result that Darwinists, in order to save face, pretend that everyone who disagrees with them is a religious crackpot (with the further result that many smart people erroneously believe that evolution itself has no real support, since they’ve only encountered it via Darwinists.) Having spent years reading Darwinian writings, alongside those of Intelligent Design theorists, Young Earth Creationists, atheist skeptics of Darwin, various kinds of spooky weirdos, and other people I don’t agree with, I’m finally in a position to explain what I think is going on. Here it is:
1) The case for evolution, considered apart from Darwinian reasoning, is fairly strong, but, by itself, suggests a teleological (purposive) evolutionary process.
2) The case for the Neo-Darwinian mechanism, considered as the main cause of evolution, is incredibly weak, but, by itself, suggests a meaningless, arbitrary process that spits out an imitation of purpose.
Since Darwinists, for philosophical reasons, — some noble; some less-so — want a blind, aimless process that also gives them order and design, they prefer option 2.
That’s it. That’s why they lead with suspect evidence, and insist on tying the cause for evolution in general to the case for the Neo-Darwinian mechanism in particular. This mechanism, or something like it, is the only sort of mechanism they want.
But there are some respectable, or at least understandable reasons for clinging to their bad mechanism. One is that the case for evolution was never purely empirical. You cannot see macro-evolution; you can only reconstruct it from the convergence of the patterns I described above. Really, we’re talking about a strong inference; a probabilistic, philosophical judgement made by surveying empirical facts. To admit this, however, makes the practical empiricist deeply uncomfortable. He feels that without an observable mechanism and a paradigm, he will be left in the realm of “mere philosophy.” Therefore, he clings to the only all-explaining mechanism he has, even though he knows it really doesn’t bear the weight he wants to place on it.
Another cause for Darwinian distress is the legitimate fear – which they often express – that admitting purpose into the process of evolution itself will open up a God-of-the-gaps problem. Essentially, the evolutionary biologist is rightly concerned about the integrity of his discipline as an independent source of knowledge. If God is always “popping in” to direct evolution, then the study of the history of life might become arbitrary. What is the point of science if, after we hit a wall of mystery, we simply have to throw up our hands and say, “a wizard did it”? Even Catholic evolutionary biologist Kenneth Miller has objected to what he sees as the notion of the Magician God. To his way of thinking, a process of creation that unfolds over time via mutation plus natural selection is more eloquent, and more worthy of the God whom Christians worship.
Now these two concerns are noble ones, and worthy of being taken seriously, but they go beyond the scope of this essay. Though I’ll deal incidentally below with the second concern, it probably merits its own future post.
Conclusion
I’d like to wrap up by asking a few loaded questions, and trying to answer them:
Q1) If you keep common descent, but remain agnostic about what exactly is providing novel genetic material, body plans, etc., then what does the overall pattern of evolution suggest about the progress of life?
A) If we just look at the progress of life from single-celled organism to the higher animals, it appears that we’re looking at process of the gradual discovery of a range of possible life forms. That is, the process itself seems somewhat groping, and subject to the whimsy of chance, but the outcomes reached seemed very non-random. There is no way that the path from land mammal to whale, or the independent production of unrelated but similar placental and marsupial mammals, could have occurred without a evolution having a structure and direction.
Q2) How could evolution be purposive and directional without being micromanaged and arbitrary (that is, without having a creator “popping in” to infuse new information into genes?)
A) By preceding within some kind of morphological framework or track, such that the groping, meandering processes of evolution (however they might actually work) are bound, given the right time and circumstances, to stumble upon (or into) very non-random paths. Just as non-living substances (water, for example) are actualized when certain atomic states are reached (because the form of water is already present in a virtual way within the potentialities of Hydrogen and Oxygen), and just as certain microscopic structures like proteins are assembled by the “happenstance” of amino acids coming together in just the right sequence so as to actualize the very specific structures that they are already ordered toward becoming (under the right circumstances), so – I suspect – it is the case that life as a whole is a series of potential structures within potential structures, forms that are virtually present in the very architecture of matter, but which only come to be as individual life forms when, according to precise sequences, the processes of nature discover them, and unfold them in ever higher cascades of kind and type. Such a process would be perfectly natural – in the sense of preceding according to ways and processes of nature – while at the same time showing the clear fingerprints of a master artist. Moreover, it would not require an inquiry-stifling God-of-the-gaps, for God’s fingerprints would be in every empirical detail, and in the pattern as a whole, while at the same time remaining appropriately elusive.
Q3) Since we don’t have evidence for “purposive” mutations, only random and stochastically isolated ones, isn’t it more scientific to say that the mutations that give rise to new life forms are random?
A) No it isn’t, for two reasons. First, because the mutations we do know about are not the kinds that can produce whole new functions or body plans, but rather the kinds that tend to do nothing, or else to cause harm. If we insist on using the word “mutation” to mean “whatever it was that happened such that reptiles began forming proto-feathers” than we are already using the word in a pre-scientific, philosophical way. Second, since the macro-changes that have occurred over vast stretches of time have clearly moved in pronounced directions toward orderly outcomes, it is much more reasonable to interpret these mysterious trajectories as purposive (teleological) even though we must, for the moment, remain ignorant of how the changes arose, and why they kept arriving just when they did. To put it another way, if we can clearly see that a speedboat, over the course of fifty million years, is gradually being changed into a submarine, it is totally unreasonable to consider this morphological direction accidental, even if – especially if – every intervening stage is functional.
Q4) Do we need to understand the mechanism of evolution to be rationally justified in believing it?
A) Contra the Darwinists and the creationists, no, we don’t. However, we need to be willing to admit that our reasons for believing in evolution in general are not purely empirical, but empirical-philosophical. Common descent (of some kind) is the best inference from the available pattern of evidence, and there are certain pieces of evidence (like retrovirus insertions) that have no viable explanation apart from common descent.
Q5) How can evolutionary biology proceed without a paradigm as robust and all-explanatory as the Neo-Darwinian synthesis, even if it has flaws?
A) By taking the true and good elements of it, and not insisting on the doubtful and discredited elements. Reasonable people do not go on forcing square pegs into round holes, and hiding discrepancies, just because this is intellectually easier. Contrary to what Neo-Darwinian dogmatists seem to believe, people would take them (and evolution) more seriously if they did not insist on knowing more about the subject than they actually do. Admitting the limits of one’s knowledge is a sign of wisdom, and of reasonability. Anyway, the object of the science of biology ought not be providing a pat origin story for everything, but rather acquiring ever deeper knowledge and wisdom about organisms, and their relations and habitats.
Q6) But wouldn’t that take us back to the days when biologists were mere stamp collectors?
A) Biologists, and all reasonable, healthy people, are mere stamp collectors! None of us knows, in advance, the whole shape of the universe. The difference between authentic wisdom and mere ideology is that wisdom is as interested in the particulars as it is in the whole, whereas ideology only values the individual fact as a means to the simple, pat, boiled-down solution, or as part of a scheme that fits inside one’s own head. The true biologist knows organisms, loves them, and is always trying to learn more. He is a collector of discrete facts. Like a stamp collector, he values variety for variety’s sake. As for schemes and plans, he has to content himself with trying to put his stamps into the correct drawers.
Q7) But who are you to say all this? You’re not a scientists!
A) No, but I’m the intelligent layperson who reads very carefully, who is the Darwinists’ presumed audience, and who should be able to follow (and be convinced by) Neo-Darwinism, if it were compelling (like in the way relativity is compelling.) I’m the guy who would rather believe the mainstream science idea, because it’s easier that way, and doesn’t give anyone cause for grouping me with Keepers Of Odd Knowledge. So, the fact that Neo-Darwinism strikes me as intellectually sloppy, even after carefully reading Darwin, Coyne, Miller, Mayer, Prothero, Gould, and others, should be cause for concern among those who promote this idea. Why are you not convincing people like me? Why do we think you’re selling us a bill of goods (at worst) or are subject to wishful, muddled thinking (at best), whereas we don’t think that about other fields in the physical sciences?
The bottom line, the whole thrust of my argument, is that the Neo-Darwinian account is full of problems, that these problems are the real reason why plenty of smart people don’t buy into it, and that Darwinists themselves do the worst damage to general belief in evolution because they insist on tying it poor arguments, inconclusive evidence, circular reasoning, and fuzzy language. Some may do so for what are partly noble concerns about the proper domains of science, religion, philosophy, and theology, while others seem to be motivated by a desire to preserve an atheist origin story about a god called Natural Selection, who bestows upon us the mere affectations of purpose and order without the troubling need to follow them upwards to their source. That is my position on the matter. But what do you think?
© 2022 Joseph Breslin All Rights Reserved